 This week we discussed the following paper:
This week we discussed the following paper:
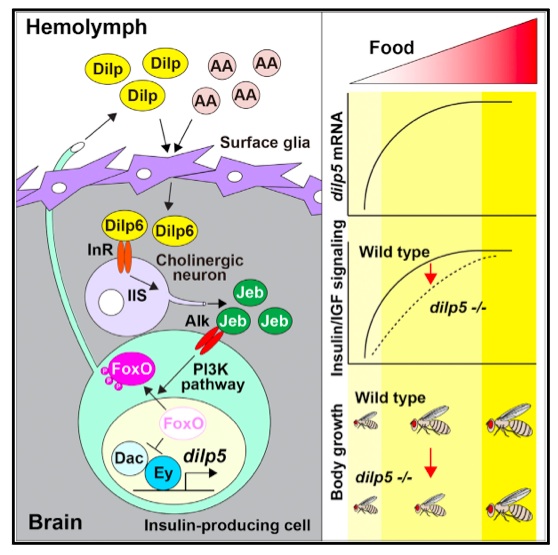
In this paper Okamoto and Nishimra describe how dILP5 mRNA expression is regulated by dietary nutrients (protein). This regulation relies on a relay mechanism involving nutrient-responsive glia, which locally secrete dILP6 to activate cholinergic neurons, which then secrete Jeb that activated the Alk receptor on the surface of IPCs, which then induce dILP5 expression in response to PI3K/Akt activation.
We liked this paper. We were interested in some of the parallels between how IPCs responded to nutrients to control dILP5 and how neuroblasts control their division in response to nutrients (in papers we have discussed in previous lab journal clubs). In both cases, an initial step is local dILP6 secretion from surface glia, however, in the case of neuroblast division, these glia respond to a fat-derived signal release in response to TOR activation (Chell, 2010; Sousa-Nunes, 2011). In this paper, activation of TOR in the fat body appears to be unnecessary. Instead the surface glia respond directly to amino acids and insulins. Similarly the Jeb-ALK pathway is also involved in controlling IPCs in neuroblasts. In this paper, ALK activation of IPCs is necessary to promote dILP5 expression in nutrient-rich conditions. In contrast ALK activation of neuroblasts is required to maintain their proliferation in late larval starvation (brain sparing – Cheng, 2011)
